Il sistema immunitario è classicamente distinto nelle sue due branche effettrici che vedono coinvolte cellule specializzate. Da un lato abbiamo la risposta immunitaria aspecifica (detta anche naturale o innata); dall’altra, l’immunità specifica (detta anche acquisita o adattativa). Molto sinteticamente, le due branche interagiscono tra loro strettamente al fine di proteggerci dai patogeni, seppure la risposta innata, guidata da cellule e molecole con attività generica (fagocitica, opsonizzante, proinfiammatoria), e quella specifica, guidata da cellule e molecole ad attività antigene-specifica (anticorpi, risposte antigene-relate), possano mantenere una teorica autonomia funzionale.
Sia la risposta innata che acquisita vengono suddivise in un braccio effettore cellulare ed in uno umorale: la risposta innata vede in gioco come elementi cellulari i granulociti (neutrofili, eosinofili, basofili), i macrofagi, le cellule dendritiche, le cellule NK e alcuni elementi della neuroglia, mentre sul piano umorale i fattori del complemento, istamina, prostaglandine, leucotrieni, le proteine della fase acuta (tra cui la proteina C reattiva); la risposta acquisita vede sul versante cellulare i linfociti T e B con le loro diverse sottopopolazioni, e sul versante umorale gli anticorpi e il vasto complesso recettoriale che ne regola la produzione. Tra i coordinatori delle interazioni e dei pattern di attivazione dei sistemi di difesa innati e acquisiti troviamo i recettori toll-like (TLRs, toll-like receptors) da cui originano gli articolati pathway capaci di indirizzare la risposta immunitaria in sensi diversi a seconda del patogeno da eliminare (batteri gram + o -, virus, tossine, miceti, parassiti, ecc…).
Si deve principalmente al lavoro di J. Edwin Blalock la dimostrazione dell’esistenza di una interfaccia che pone in comunicazione i sistemi nervoso ed endocrino con quello immunitario. L’interfaccia è costituita dalla capacità di pressoché tutti gli elementi costitutivi dei tre sistemi di produrre molecole e sostanze condivise e di riconoscerne la presenza tramite specifici recettori.
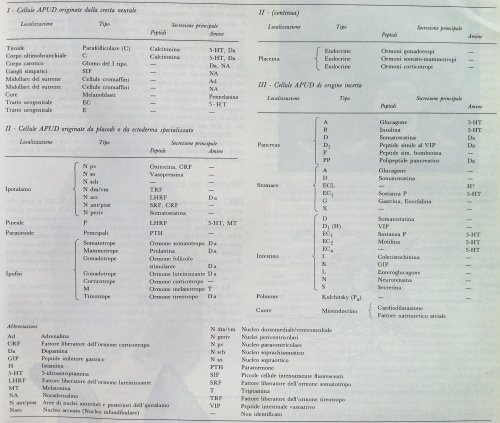
Nella figura al presente link viene schematizzata la grande capacità sintetizzante e recettoriale dei linfociti T. I linfociti, cellule cardine della risposta acquisita, da semplici cellule immunitarie diventavano nel corso delle ricerche anche cellule a forte impronta endocrinologica, una rivoluzione in immunologia. Lo scambio di segnali, grazie alla capacità di sintesi delle cellule immunitarie, si presenta come reciproco: non si tratta solo di centri superiori (ad es. la regolazione neuroendocrina ipotalamica) che modulano centri effettori (la risposta immunitaria) ma anche gli effettori sono in grado di produrre molecole con azione stimolatoria o inibitoria sui centri neuroendocrini. La Tabella 1 (Table 1), tratta dall’articolo di JE Blalock “The immune system as the sixth sense” Journal of Internal Medicine 2005; 257: 126–138, mostra come i principali mediatori ormonali e i neurotrasmettitori vengano secreti dalle cellule immunitarie.
Bisogna tuttavia precisare come gli studi atti a dimostrare la capacità di produrre determinate molecole siano principalmente in vitro. Questo ha da un lato il vantaggio di rendere i risultati più specifici per le cellule in esame ma dall’altro il limite di non poter determinare la reale portata in vivo della produzione di tali molecole. In altre parole, quale è veramente l’impatto nell’essere umano di tale sintesi in tali cellule? E’ significativo o è trascurabile? Dobbiamo tenere presente infatti che tutte le cellule del nostro organismo hanno lo stesso corredo cromosomico e gli stessi geni (fatta eccezione per le cellule sessuali, ossia spermatozoi e ovociti): ciò che le differenzia non sono i geni ma la loro diversa espressione nei diversi organi e tessuti. In linea teorica qualsiasi cellula del nostro organismo potrebbe essere indotta a sintetizzare qualsiasi sostanza prodotta fisiologicamente da un’altra cellula.
Fatta questa precisazione, procediamo col vedere le interazioni più frequenti e, per così dire, più consistenti a livello dell’interfaccia neuro-endocrino-immunitaria. Per primo, c’è uno stretto circuito regolatorio mediato dall’asse ipotalamo-ipofisi-surrene: la modulazione che si verifica reciprocamente tra SNC e immunità è a questo livello fondamentale. Stress, fatica, restrizione calorica, deprivazione di sonno, sforzi ecc… attivano la produzione di CRH ipotalamico con conseguente incremento dell'ACTH e dei valori di cortisolo circolanti. Le cellule immunitarie (neutrofili, macrofagi, linfociti) possiedono recettori per il cortisolo che portano in caso di stimolo protratto a immunosoppressione con ritiro dei neutrofili dai tessuti (reclutamento nel torrente ematico e conseguente aumento dei loro livelli plasmatici), azione antilinfocitaria sia per inibizione diretta che per inibizione della produzione di numerose citochine, inibizione della reazione infiammatoria tramite blocco della sintesi delle prostaglandine. In particolare, gli effetti immunosoppressivi più spiccati si osservano sulla popolazione T linfocitaria, monocitaria e cellulare dendritica con:
- inibizione dell’espressione endoteliale di molecole di adesione quali ELAM-1, ICAM-1, E-selectina e del CD18 (meccanismi in gioco anche nella centralizzazione nel torrente ematico dei neutrofili e loro inibizione nel raggiungere i tessuti infiammati, della riduzione numerica dei monociti e ridistribuzione in generale nei siti di deposito quale il midollo osseo);
- inibizione della secrezione delle citochine macrofagiche quali IL-1, IL-6, IL-8, MCP-1, TNF-alfa e delle linfochine del profilo Th1 come IL-2, IL-12, IFN-gamma, e ancora TNF-alfa;
- blocco dell’ espressione di antigeni HLA-II sui macrofagi, sulle cellule di Langerhans e sulle cellule dendritiche;
- inibizione della sintesi delle proteine complementari e induzione dell’apoptosi negli eosinofili e nei linfociti T.
Da questa breve rassegna sugli effetti immunologici mediati dal cortisolo e, quindi, controllati dal circuito a retroazione costituito dall’asse ipotalamo-ipofisi-surrene, possiamo intuire la fondamentale connessione tra sistema endocrino e immunitario. Inoltre, essendo le stesse cellule immunitarie (in particolare linfociti T, B e i macrofagi) in grado di produrre corticotropina (ACTH), lo stesso sistema immunitario è in grado di intervenire nella regolazione ormonale modulando la sintesi del cortisolo stesso e la produzione di CRH. Le cellule immunitarie presenti nella corticale dei surreni (linfociti, cellule dendritiche, mastcellule, macrofagi) una volta attivate sono altresì in grado di produrre citochine con diversi effetti sulla secrezione glucocorticoide: IL-1 e IL-6 sembrano possedere effetti stimolatori sulla secrezione di cortisolo mentre il TNF-alfa sembra al contrario possedere effetti inibitori.
L’interfaccia neuroendocrino-immunitaria vede in primo piano anche la midollare del surrene ed in generale il sistema nervoso adrenergico. Pressoché tutte le cellule immunitarie sia a livello del braccio innato che acquisito dell’immunità sono dotate di recettori per adrenalina e noradrenalina, in particolare di beta-recettori (più frequentemente beta-2). Recettori alfa-adrenergici sono presenti sulle cellule dendritiche e monocitarie, mentre beta-recettori sono presenti sui neutrofili, sui monociti/macrofagi, sulle cellule dendritiche, sulle cellule NK ed in molte popolazioni T e B linfocitarie. In generale, la stimolazione adrenergica acuta in vivo produce un aumento numerico delle cellule NK circolanti mentre sembra ridurre l’attività dei neutrofili con conseguente azione antinfiammatoria. Questa sembra esplicarsi anche a livello delle cellule dendritiche e della linea monocitomacrofagica. Il ruolo del sistema adrenergico a livello delle popolazioni T e B sembra più complesso, dove è stata riscontrata sia stimolazione che inibizione dell’attività cellulare CD4+ e/o CD8+ a seconda dello stimolo antigenico usato ed in relazione anche alle interazioni con i linfociti B. Infatti, il tono adrenergico al momento del primo contatto con l’antigene sembra fondamentale per il successivo sviluppo della risposta anticorpale. Lo stimolo adrenergico si correla positivamente con più alti titoli anticorpali nella risposta primaria all’antigene mentre scarso sembra l’effetto del tono adrenergico nei successivi contatti con l’antigene. Tuttavia è interessante notare che la somministrazione di agenti beta-2-adrenergici insieme a cortisolo aumenta la sintesi di IgG sieriche nei soggetti asmatici permettendo di ipotizzare un contributo adrenocortisolico a livello dello switch isotipico.
Ci pare importante ora evidenziare che, nell’interpretazione degli effetti adreno-mediati sul sistema immunitario, grande importanza ha l’espressione recettoriale cellulare, che varia notevolmente a seconda dello stato (naive, quiescente, attivata, iperattivata) della cellula immunitaria in esame. Ad esempio, uno stimolo noradrenergico su cellule NK coltivate con IL-2 mostravano una ridotta espressione di IFN-gamma e TNF-alfa con conseguente effetto inibitorio sulla loro attività. L’ IL-2 viene prodotta principalmente dai linfociti T attivati, quindi in una situazione di risposta immunitaria in atto.
Provando a trarre delle conclusioni generali, assai diverso può essere l’effetto della stimolazione adrenergica al di fuori di un sistema immunitario attivato, durante la sua attivazione o dopo di essa, rispettivamente con effetti stimolatori nel primo caso, inibitori nel secondo e trascurabili nel terzo. Altro dato importante da tenere presente, l’attivazione immunitaria può essere sistemica (definita anche come SIRS, systemic inflammatory response syndrome, cioè una sindrome da risposta infiammatoria generalizzata che si caratterizza generalmente per la presenza di febbre, leucocitosi, tachicardia, tachipnea) o localizzata (infezioni localizzate, traumi limitati) dando ragione di effetti diversi su popolazioni cellulari nello stesso individuo ma in contesti tessutali specifici.
Anche la stimolazione colinergica regola l’immunità così come quest’ultima interviene nella regolazione della bilancia simpato-vagale. Il nervo vago costituisce un importante sistema antinfiammatorio colinergico capace di intervenire sia nell’infiammazione sistemica (ad es. in corso di endotossinemia) sia localizzata. L’acetilcolina si lega alla subunità alfa-7 del recettore nicotinico espressa sui macrofagi inibendone il rilascio delle citochine proinfiammatorie quali TNF-alfa, IL-1, Il-6, IL-18, HMGB1 senza interferire con la produzione di citochine antinfiammatorie quale IL-10, modulando quindi potentemente la risposta infiammatoria. I recettori nicotinici sono altresì presenti sulle cellule endoteliali, dove lo stimolo colinergico è in gradi di indurre ipoespressione delle molecole di adesione per diversi leucociti, soprattutto i neutrofili, riducendo quindi la loro migrazione con conseguente limitazione della flogosi. L’ipertono vagale è tuttavia talvolta associato alle sindromi respiratorie broncocostrittive quali asma e broncopneumopatia cronica ostruttiva. In questo caso tuttavia l’azione è mediata a livello dei recettori muscolari bronchiali muscarinici, i quali inducono contrazione della muscolatura liscia bronchiale. I bloccanti prevalentemente muscarinici, quali tiotropio, oxitropio e ipratropio sono in grado di esercitare un effetto antimuscarinico cercando di mantenere i benefici dell’ipertono vagale sui recettori nicotinici.
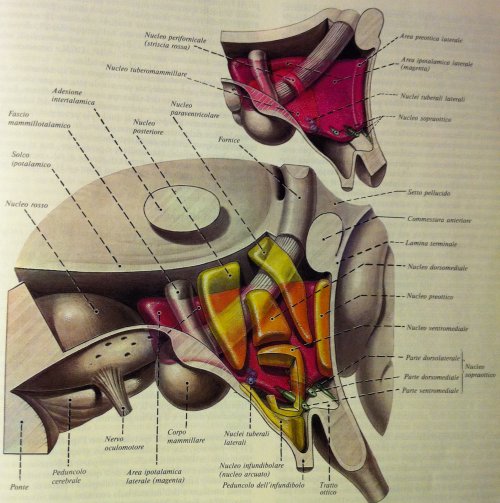
I linfociti sono a loro volta in grado di produrre acetilcolina, intervenendo peraltro nella modulazione infiammatoria colinergica, così come le cellule timiche (timociti) e spleniche (splenociti) sono capaci di produrre in quantità significativa adrenalina; ciò rende il sistema immunitario stesso parte integrante della regolazione simpato-vagale, ossia del sistema nervoso autonomo. Inoltre il timo, organo cardine nell’ontogenesi dei linfociti T e soggetto a progressiva involuzione nell’età adulta, produce diversi fattori peptidici denominati qui genericamente timosine, in grado di regolare la sintesi di ACTH e beta-endorfina a livello diretto ipofisario, così come di gonadorelina a livello ipotalamico la quale agisce sulla secrezione di gonodatropine ipofisarie e quindi sulla maturazione sessuale sia nel maschio che nella femmina.
Gli estrogeni, come ben noto da parecchi anni, sono capaci di intensificare l’attività linfocitaria rendendo ragione della maggior prevalenza di malattie autoimmunitarie nel sesso femminile. Così anche GH e prolattina sono in grado di stimolare la proliferazione linfocitaria attraverso recettori di membrana specifici. Gli stessi linfociti sono in grado di sintetizzare GH e prolattina modulando teoricamente non solo per via autocrina e paracrina il proprio trofismo, ma anche i circuiti di feedback retroattivo ipotalamo-ipofisari dando spiegazione, in sinergia con altri fattori, ai deficit del profilo costituzionale somatico degli individui affetti da patologie infiammatorie croniche, specie se originatesi in epoca precoce quale l’età evolutiva.
I linfociti sono anche in grado di produrre TSH, intervenendo nei processi funzionali legati all’asse ipotalamo-ipofisi-tiroideo, mentre la linea cellulare monocitomacrofagica è in grado di sintetizzare in maniera significativa molecole ad azione pleiotropica quali il polipeptide intestinale vasoattivo (VIP) e la somatostatina, due molecole ad attività ormonale complessa agenti sia sull’apparato digerente (ad es. sulla funzionalità pancreatica) che sul sistema nervoso come neuromodulatori centrali e periferici (sistema nervoso enterico).
A scopo riassuntivo riportiamo alcune tabelle tratte principalmente dalla bella revisione PNEI nell’ambito del Trattato Italiano di Medicina di Laboratorio fondato da Angelo Burlina ed edito da Piccin (Vol II/2).
Principali influenze del sistema neuroendocrino su quello immunitario.
Effetti delle citochine sull’asse ipotalamo-ipofisi-surrene.
Effetti delle citochine sull’asse ipotalamo-ipofisi-gonadi.
Effetti delle citochine sull’asse ipotalamo-ipofisi-tiroide.
L’interfaccia endocrino-immunitaria vede anche in azione la vitamina D3 che, per certi aspetti, ha caratteristiche assai vicine al concetto di ormone (mediatore intercellulare sistemico) più che di vitamina (ossia cofattore necessario allo svolgimento di reazioni enzimatiche). Infatti essa interviene non solo nel metabolismo del calcio ma anche nella regolazione del sistema immunitario:
- a livello delle cellule dendritiche inibisce l’espressione dei complessi peptide/MHC II modulando quindi la presentazione dell’antigene in senso inibitorio;
- ancora a livello delle cellule dendritiche, inibisce la produzione di IL-12 con la conseguente induzione di una risposta Th2 invece che Th1;
- inibizione T linfocitaria diretta sulla produzione delle citochine Th1 quali IL-2 e IFN-gamma;
- induzione dei linfociti T-reg.
E’ necessario comunque precisare che in vivo le concentrazioni di vitamina D3 richieste per la modulazione del sistema immunitario sono generalmente maggiori di quelle fisiologiche circolanti e quindi solo raggiungibili per valori sovrafisiologici che si correlano a iperaccumulo di calcio.
Per concludere, senza pretesa di esaustività di un argomento così complesso e articolato, il sistema neuroendocrino ed il sistema immunitario sono intimamente connessi e reciprocamente influenzati a livello di una vastissima interfaccia costituita da recettori e molecole capaci di mantenere l’omeostasi dell’organismo. A loro volta tali molecole sono in grado di influenzare direttamente l’eccitazione o l’inibizione neuronale attraverso diversi meccanismi che vedono in gioco circuiti elettrici e biochimici tra tutti quegli elementi cellulari che vanno a costituire il sistema nervoso centrale (SNC) ossia, oltre ai neuroni, anche le cellule gliali, le cellule neuro-endoteliali, le cellule neuro-endocrine e le cellule neuro-immunitarie (ossia quelle cellule immunitarie periferiche che per diversi motivi fisiopatologici sono migrate a livello del SNC). In tal senso anche il sistema nervoso autonomo gioca un ruolo essenziale: grazie alla sua sensibilità alle variazioni cellulari e umorali che avvengono in periferia è capace di veicolare tali variazioni fino ai centri diencefalici e telencefalici. L’ipotalamo in particolare funziona come un filtro che lascia passare meno dell’1% delle informazioni che giungono dalla periferia verso la corteccia cerebrale. Il SNC può venire comunque informato sull’attivazione del sistema immunitario sia attraverso la via umorale (endocrina e citochinica) sia attraverso le vie nervose ascendenti. Questa è la base fisiopatologica delle risposte centrali (febbre, anoressia, astenia, iperalgesia, cefalea, apatia, deflessione del tono dell’umore, ecc…) agli stati di malattia. Questa costituisce la base della risposta cognitivo-comportamentale al sistema immunitario: l’interfaccia psico-immunitaria.